График работы:
E-mail:
Бесплатная консультация
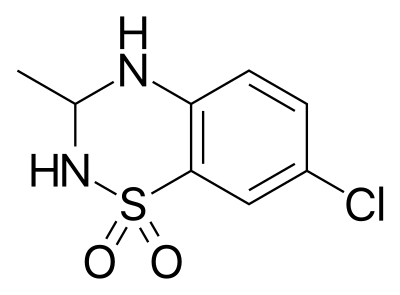
| Номер CAS | 22503-72-6 |
| Другие имена | IDRA 21, 22503-72-6, 7-хлор-3-метил-3,4-дигидро-2H-1,2,4-бензотиадиазин 1,1-диоксид, 689UW7PT68, (+)-IDRA-21, (-)-IDRA-21, Biomol-NT_000225, D0S0VZ |
| Название по IUPAC | 7-хлор-3-метил-3,4-дигидро-2H-1λ6,2,4-бензотиадиазин 1,1-диоксид |
| Молекулярная формула | C₈H₉ClN₂O₂S |
| Молекулярный вес | 232.69 |
| Хранение | Хранить в сухом прохладном месте, вдали от прямых солнечных лучей. |
IDRA 21 (7-хлоро-3-метил-3-4-дигидро-2H-1,2,4 бензотиадиазин S,S-диоксид) является ноотропным соединением, которое действует как аллостерический модулятор рецепторов глутамата AMPA. Ноотропные агенты продемонстрировали потенциал в улучшении познавательных способностей посредством ингибирования скорости десенсибилизации рецепторов AMPA. При этом IDRA 21 полагается на высвобождение эндогенного глутамата для укрепления глутаминергических синапсов. Текущие исследования в отношении ноотропов показывают, что это соединение обладает способностью обращать вспять эффекты амнезии до лечения, усиливать отсроченное обучение и ослаблять вызванные дефициты обучения. [1]
1) Результаты исследования подтверждают эффективность использования модуляторов AMPA, таких как IDRA 21, в случаях нарушения памяти или когнитивных функций.
2) Исследование показало, что IDRA 21 обладает потенциалом для снижения фармакологически вызванных когнитивных нарушений.
1) Исследовательская группа Buccafusco et al изучает способность IDRA 21 ингибировать индуцированную глутаматом инактивацию мембранных токов в клетках эмбриональной почки человека (HEK) в зависимости от концентрации. Электрофизиологические исследования начались со стабильной трансфекции рекомбинантных клеток HEK 293 с группой человеческих рецепторов GLuR ½ flip в DMEM и 10% сыворотки плода теленка.
Внутриклеточная среда, используемая в этой части исследования, состояла из NaCL 15 мМ, тетраэтиламмоний-Cl 20 мМ, CsF 110 мМ, 1,2-бис-(2-аминофенокси)-этан-тетрауксусной кислоты (BAPTA) 11 мМ, MgCl2 2 мМ, CaCl2 1 мМ, 1,4 (2-гидроксиэтил)-пиперазин-1-этансульфоновой кислоты (EHPES) 10 мМ и АТФ 1 мМ при pH 7,2. С другой стороны, внеклеточная среда, которую использовали, состояла из NaCL 140 мМ, KCl 5,3 мМ, CaCl2 1,8 мМ, MgCl2 0,8 мМ, глюкозы 21,5 мМ, 1,4-(2-гидроксиэтил)-пиперазин-1-этансульфоновой кислоты (HEPES) 10 мМ при pH 7,4. Кроме того, и глутамат, и IDRA 21 применялись с использованием пьезоприводной системы проточной трубы, однако ноотроп предварительно применял в течение 30-60 секунд в отсутствие глутамата для достижения устойчивого состояния эффектов на агонистические реакции [1].
Это исследование также включало животную модель из четырнадцати макак-резусов, которым вводили IDRA 21 и непрерывно оценивали с помощью задачи отложенного сопоставления с образцом (DMTS). Ноотроп вводили обезьянам, заставляя исследователей взвешивать соответствующее количество каждый день с точностью до 0,1 мг. Затем взвешенный ноотроп смешивали с 20 мл напитка для приматов со вкусом фруктового пунша, и либо активная доза, либо растворитель вручную вводились через шприц за 1 час до тестирования DMTS. Хотя животные проходили тестирование DMTS каждый будний день, между каждым введением IDRA 21 или раствора проходило 5 дней [1].
Обезьянам был предоставлен неограниченный доступ к водопроводной воде, и они питались стандартным лабораторным кормом для обезьян, дополненным различными фруктами и овощами. График кормления позволял получать около 15% ежедневного потребления пищи из 300 мг гранул подкрепляющей пищи. Исследователи также отметили, что каждый испытуемый ранее принимал участие в одном или нескольких обратимых фармакологических исследованиях лекарственных препаратов и им был разрешен 4-недельный период вымывания перед введением IDRA 21 [1].
Животных тестировали в их индивидуальных домашних клетках с использованием автоматизированной компьютерной системы обучения и тестирования, предназначенной для измерения и категоризации задержек и латентности реакции при каждой проблеме сопоставления. Каждая ежедневная сессия включала 96 испытаний; испытание начиналось, когда один из трех цветных дисков загорался и нажимался обученной обезьяной. Аналогичное действие прекращало освещение клавиши образца во время интервала задержки. После интервала задержки загорались две тестовые клавиши, а не клавиша образца. Одна из двух клавиш была того же цвета, что и клавиша образца, и целью обезьяны было выбрать диск того же цвета, что и клавиша образца. Если им удавалось сопоставить, то испытуемый получал вознаграждение за правильный ответ [1].
Каждое испытание было разделено на 5-секундный интервал, и исследовательская группа приняла к сведению несколько мер предосторожности, включенных в исследование, включая: различные комбинации цвета стимула, сколько раз появлялся каждый цвет и каждая цветовая комбинация, противовес между левым и правым и показанными цветами, а также процедуры противовеса стимула и то, как они были сопоставлены с длиной задержки интервала. При этом цвета стимула (красный, желтый и зеленый) были расположены таким образом, чтобы каждый цвет появлялся равное количество раз в качестве образца, и каждый цвет появлялся равное количество раз в качестве клавиш выбора. Кроме того, когда два цвета появлялись в комбинации, исследователи меняли их левое/правое расположение на непредсказуемый шаблон. [1]
Ежедневные испытания, которые проходила каждая обезьяна, были представлены с использованием трех различных, но фиксированных последовательностей рандомизированных интервалов задержки и цветов выбора, которые чередовались ежедневно. Эта процедура снижала вероятность того, что животные запомнят первые несколько испытаний в сеансе. На индивидуальном уровне обезьяны обладают способностью поддерживать производительность сопоставления после различного времени удержания; самая длинная задержка, выбранная для обезьяны, позволяла последовательно правильно сопоставлять выше уровня случайности. Длина интервала задержки корректировалась для каждой обезьяны до тех пор, пока не были определены различные уровни сложности выполнения. Эти уровни включали: 1) наименее сложную нулевую задержку; 2) короткий интервал задержки; 3) средний интервал задержки; и 4) длинный интервал задержки, представляющий предел каждой обезьяны с точки зрения их производительности во время тестирования DMTS [1].
Процент правильных ответов в испытаниях с нулевым, коротким, средним и длинным интервалами задержки и тремя различными задержками выполнения задания регистрировался для всех 96 испытаний каждый день. Три задержки выполнения задания регистрировались и включали: 1) задержки выполнения образца, определенные как временной интервал между предъявлением клавиши образца и нажатием испытуемым клавиши образца для начала испытания; 2) задержки выбора во время испытаний с правильным ответом, определенные как временной интервал между предъявлением стимулов выбора и нажатием испытуемым клавиши выбора; и 3) задержки выбора во время испытаний с неправильным ответом. Исходные данные анализировались с использованием многофакторного дисперсионного анализа с повторными измерениями, в то время как для сравнения отдельных средних значений, полученных от животных во время тестирования DMTS, использовался ортогональный t-тест наименьших квадратов средних [1].
2) Исследовательская группа Томпсона и соавторов изучала влияние IDRA 21 на когнитивные нарушения, вызванные фармакологически у обезьян патас. Для целей данного исследования в качестве испытуемых выступали две самки и два самца обезьян патас. Каждый испытуемый имел опыт использования поведенческих процедур и имел предыдущее воздействие обратимых экспериментальных препаратов. При этом испытуемые не принимали препараты в течение как минимум 2 недель до начала исследования. Вода была постоянно доступна всем испытуемым, и животные поддерживались на уровне примерно 90% от своего веса при свободном кормлении с помощью диеты из гранул с банановым вкусом, обезьяньего корма и различных фруктовых и витаминных добавок [2].
Обезьян лечили как алпразоламом, так и IDRA 21. Алпразолам вводили перорально, два раза в неделю в дозах от 0,01 до 0,32 мг/кг. Пероральный раствор готовили путем суспендирования соединения в 5% растворе 2-гидроксипропил-бета-циклодекстрина, смешанном с 15 мл фруктового пунша, который испытуемый пил каждый вторник и пятницу. Затем обезьянам предварительно давали рацемическую смесь IDRA 21 в дозах от 0,3 до 5,6 мг/кг. Ноотроп суспендировали в Tween80 и давали испытуемым таким же образом, как и алпразолам. Каждое лечение тестировали отдельно и в сочетании друг с другом с воздействием IDRA 21 в течение 120 минут и воздействием алпразолама в течение 60 минут. Кроме того, от 1 до 30 мг/кг анирацетама вводили субъектам для прямого сравнения с IDRA 21. Каждый субъект был размещен в стандартной, специфичной для приматов индивидуальной клетке, которая была оборудована панелями ответа для отслеживания активности во время поведенческого тестирования. Компонент получения теста начинался с проецирования одной из четырех геометрических форм на красный фон на трех различных ключах ответа [2].
Целью теста было выучить цепочку из четырех ответов, нажимая правильную клавишу в присутствии каждой формы. Формы включают в себя: горизонтальную линию, левое правильное, треугольник, правое правильное, вертикальную линию, центральное правильное, круг и правое правильное. После завершения цепочки из четырех ответов свет выключался, и загоралась контрольная лампа возле дозатора гранул. Цепь сбрасывалась после нажатия контрольной лампы, и устанавливалось фиксированное соотношение, которое планировало подачу пищи после каждого пятого завершения цепочки и нажатия контрольной лампы, чтобы поддерживать цепочку из четырех ответов. Для сравнения, когда один из испытуемых нажимал неправильную клавишу, наступал 5-секундный тайм-аут, во время которого клавиши были темными, а все ответы были неэффективными. При этом ошибка не сбрасывала цепочку, и стимул оставался таким же, как и до 5-секундного тайм-аута. Во время фазы освоения цепочки из четырех ответов менялись от сеанса к сеансу, и каждая цепочка выбиралась одинаковой несколькими способами.
Во время компонента производительности поведенческого теста четыре геометрические формы проецировались на зеленый фон, в то время как рисунок лево-центр-лево-право цепи из четырех ответов оставался неизменным на протяжении всех сессий. Во всех других аспектах процедуры компонента производительности поведенческого теста были идентичны процедурам компонента приобретения поведенческого теста. Сессии проводились ежедневно. Каждое испытание начиналось с периода приобретения в соответствии с процедурами, описанными выше, за которым следовал компонент производительности. Испытуемые переходили к компоненту производительности после 10 подкреплений производства пищи или через 15 минут. Каждый сеанс заканчивался через 2 часа или после того, как было достигнуто 60 подкреплений для самок и 100 подкреплений для самцов [2].
1) Результаты электрофизиологической части исследования показали изменения в мембранных токах, опосредованных глутаматом, в ответ на введение IDRA21. Обычно при нанесении 1 мМ глутамата на клетки, экспрессирующие рецепторы GLuR ½ флип, токи быстро инактивируются. Однако при нанесении IDRA 21 было показано, что постоянная времени инактивации увеличивается по мере увеличения концентрации ноотропа. Общий перенос заряда в течение первых 50 мс нанесения глутамата использовался в качестве сравнительной меры для наблюдаемых эффектов IDRA 21. Было обнаружено, что перенос заряда удваивается при концентрации 70 мкМ, что указывает на то, что применение ноотропа может способствовать возбуждающей нейротрансмиссии через рецепторы GluR ½ флип [1].
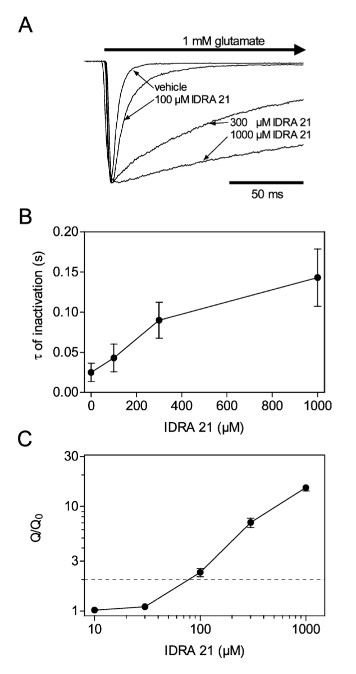
Рисунок 1: (A) Изменения в мембранном токе, вызванные глутаматом в клетках, экспрессирующих рекомбинантные человеческие рецепторы GluR ½ flip. (B) Эффекты IDRA 21 на инактивацию ответа глутамата. Время инактивации увеличивалось в зависимости от концентрации. (C) Изменения в общем переносе заряда в ответ на аппликацию IDRA 21 в течение первых 50 мс аппликации глутамата.
Что касается поведенческих исследований, проведенных на макаках-резусах, результаты были классифицированы на молодых и пожилых субъектов. IDRA 21 вводился молодым субъектам в последовательности дозирования, начиная с 0,3 мг/кг, продолжая 1,0, 3,0, 6,0, 0,15 и 10,0 мг/кг. Животные проходили тестирование через 1, 24 и 48 часов после введения препарата; испытуемым не давали ни носителя, ни ноотропа до тестирования в течение 24- и 48-часовых сеансов испытаний. Статистический анализ показал, что существует значительная связь между дозой препарата и днем тестирования, представленная двухфазным улучшением точности выполнения задач. Эти результаты были получены в сеансах со средним и длинным интервалом задержки, при этом самая низкая точка в связи наблюдалась при средних дозах IDRA 21.
Для испытаний, которые проводились через 1 час после введения IDRA 21, наблюдалась статистически значимая связь между общим эффектом лечения препаратом и производительностью во время испытания. Постанализ показал, что дозы лечения 0,15 мг/кг, 0,3 мг/кг и 6 мг/кг вызывали наиболее заметные изменения по сравнению с животными, которым вводили растворитель. Кроме того, для испытаний, которые проводились через 24 часа после введения IDRA 21, дозы, которые привели к наиболее значительным различиям по сравнению с лечением растворителем, составляли 0,15 мг/кг, 0,3 мг/кг, 6 мг/кг и 10 мг/кг. Наконец, для испытаний, которые проводились через 48 часов после введения ноотропа, дозы, которые привели к наиболее значительным различиям по сравнению с лечением растворителем, составляли 0,15 мг/кг, 3 мг/кг и 10 мг/кг. Эти результаты позволили исследовательской группе сделать вывод, что введение IDRA 21 повышает точность выполнения задач у молодых обезьян на устойчивом уровне в течение как минимум 48 часов после введения [1].
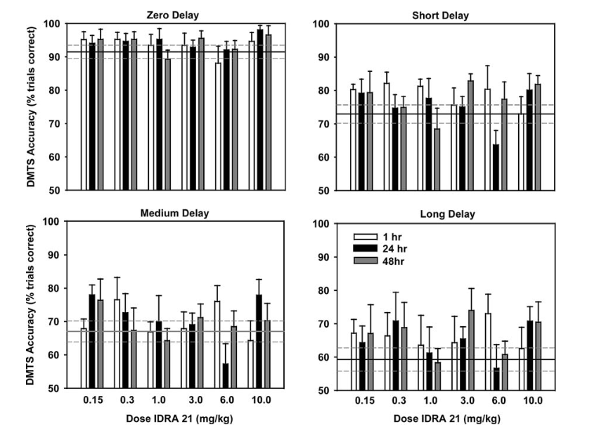
Рисунок 2: Влияние IDRA 21, вводимого в дозах 0,15, 0,3, 1,0, 3,0, 6,0 и 10 мг/кг, на точность выполнения задания через 1 час, 24 часа и 48 часов после введения ноотропа у молодых обезьян, прошедших тест DMTS.
Что касается пожилых испытуемых, каждое животное было более чем на два года старше молодой группы испытуемых. Возрастное ухудшение задач также принималось во внимание в старшей группе, и продолжительность интервалов задержки была сокращена почти вдвое по сравнению с младшей группой. Исследовательская группа также посчитала важным отметить, что молодая группа состояла из всех самцов, в то время как пожилая группа включала 4 самцов и 3 самок. Роль пола не принималась во внимание в ходе этого исследования, однако ни одна из самок не была зарегистрирована в эструсе, и не было замечено никаких очевидных эффектов, связанных с полом, во время выполнения ими задач.
Подобно более молодой группе обезьян, IDRA 21 вводился пожилым субъектам в последовательности дозирования 0,3, 1,0, 30,0, 6,0, 0,15 и 10,0 мг/кг с последующим тестированием через 1 час, 24 часа и 48 часов после введения ноотропа. Перед 24- и 48-часовыми сеансами тестирования животным не вводили ни лекарство, ни носитель. Статистический анализ результатов показал, что существует значительный эффект между дозой препарата в зависимости от интервала задержки. Также была заметная картина улучшения точности выполнения задач во время испытаний, связанных со средними и длинными интервалами задержки. Исследователи сравнили с более молодой группой субъектов и отметили, что у пожилых субъектов наблюдалась похожая двухфазная природа дозовых соотношений, однако в этой группе она была не столь очевидна [1].
Для испытаний, проведенных через 1 час после введения IDRA 21, наблюдалась тенденция к эффекту ноотропного лечения, который не зависел от дозировки и интервала задержки. Дальнейший анализ показал, что эти эффекты были наиболее значимо связаны с дозой 0,15 мг/кг и 10 мг/кг. Кроме того, для испытаний, проведенных через 24 часа после введения IDRA 21, наблюдался значительный эффект ноотропного лечения, связанный с дозами 1 мг/кг и 10 мг/кг. Наконец, для испытаний, проведенных через 48 часов после введения IDRA 21, не было никаких значительных эффектов, вызванных ноотропом. Исследовательская группа отметила это как заметное различие между более молодыми и пожилыми группами испытуемых [1].
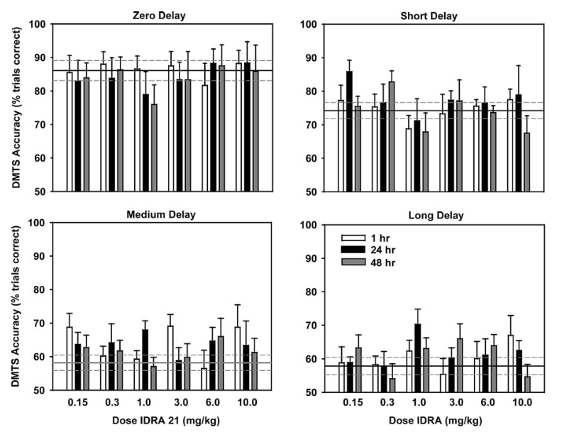
Рисунок 3: Влияние IDRA 21, вводимого в дозах 0,15, 0,3, 1,0, 3,0, 6,0 и 10 мг/кг, на точность выполнения задания через 1 час, 24 часа и 48 часов после введения ноотропа у старых обезьян, прошедших тест DMTS.
2) Результаты исследования, проведенного Томпсоном и соавторами, изначально показали, что существуют индивидуальные различия в показателях контрольного ответа как в компонентах приобретения, так и в компонентах выполнения поведенческого тестирования. Когда алпразолам вводился в дозах 0,32 мг/кг для самок и 0,1 мг/кг для самцов, показатели ответа в целом снижались в обоих компонентах, однако снижение было более заметным в период приобретения. Было обнаружено, что предварительное лечение с помощью IDRA 21 противодействует эффекту увеличения ошибок алпразолама дозозависимым образом как в компонентах приобретения, так и в компонентах выполнения поведенческого тестирования. Самая низкая доза IDRA 21, которая, как было показано, повлияла на эффект снижения скорости алпразолама, составляла либо 0,3 мг/кг, либо 1 мг/кг в зависимости от того, был ли субъект мужчиной или женщиной [2].
В то время как предварительная обработка IDRA 21 снижала эффекты алпразолама, увеличивающие ошибки, введение ноотропа в одиночку не приводило к каким-либо значительным изменениям в скорости измерения ни в одном из компонентов поведенческого теста. Это было похоже на анирацетам, исходное соединение IDRA 21, которое не вызывало никаких изменений в скорости или точности при введении само по себе. Также похоже на IDRA 21, было обнаружено, что анирацетам противодействует эффектам алпразолама, снижающим скорость, в зависимости от дозы. Однако важно отметить, что было обнаружено, что анирацетам примерно в 10 раз менее эффективен, чем IDRA 21. Ослабление снижения скорости при дозе анирацетама 30 мг/кг было эквивалентно тому, что наблюдалось при дозе IDRA 21 3 мг/кг [2].
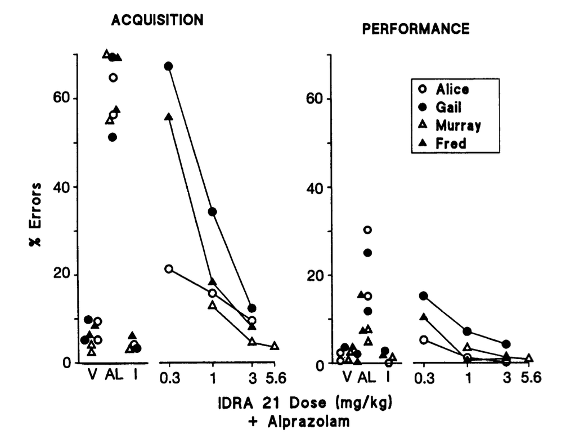
Рисунок 4: Влияние IDRA 21 в сочетании с алпразоламом на точность и ошибки в компонентах освоения и выполнения проведенного поведенческого теста.
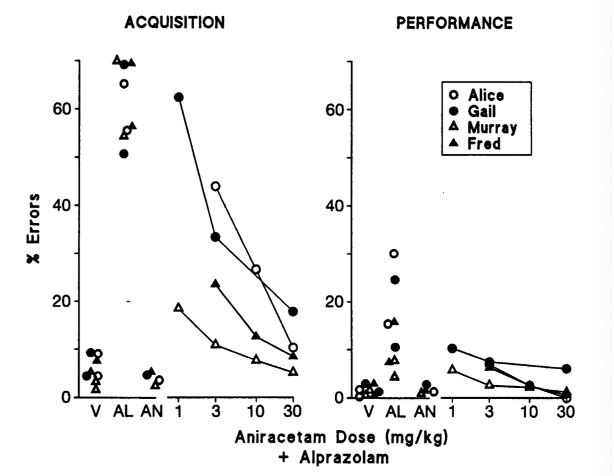
Рисунок 5: Влияние анирацетама в сочетании с алпразоламом на точность и ошибки в компонентах освоения и выполнения проведенного поведенческого теста.
[1] Buccafusco, Jerry J., Thomas Weiser, Karin Winter, Klaus Klinder and Alvin V. Terry. “The effects of IDRA 21, a positive modulator of the AMPA receptor, on delayed matching performance by young and aged rhesus monkeys.” Neuropharmacology 46 (2004): 10-22.
[2] Thompson DM, Guidotti A, DiBella M, Costa E. 7-Chloro-3-methyl-3,4-dihydro-2H-1,2,4-benzothiadiazine S,S-dioxide (IDRA 21), a congener of aniracetam, potently abates pharmacologically induced cognitive impairments in patas monkeys. Proc Natl Acad Sci U S A. 1995 Aug 15;92(17):7667-71. doi: 10.1073/pnas.92.17.7667. PMID: 7644474; PMCID: PMC41206.
© 2026 АнкеБио. Magazin nootropov Biohaker
127299, г. Москва, ул. Шоссе Энтузиастов, 56 стр. 32
+7 (495) 227-22-05
+7 (985) 227-22-05
Email: ankebiorus@gmail.com
Вы нашли для себя препарат, который хотите
приобрести по низкой цене и с гарантией
безопасной доставки?
Звоните нам
Мы Вам обязательно ответим!