График работы:
E-mail:
Бесплатная консультация
| Номер CAS | 1401708-83-5 |
| Другие имена | L-изолейцинамид, N-(1-оксогексил)-L-тирозил-N-(6-амино-6-оксогексил)-; 9WYX65A5C2; N-гексановый-Тир-Иле-(6) амид аминогексановой кислоты; PNB-0408 |
| Название по IUPAC | (2S,3S)-N-(6-амино-6-оксогексил)-2-[[(2S)-2-(гексаноиламино)-3-(4-гидроксифенил)пропаноил]амино]-3-метилпентанамид |
| Молекулярная формула | C₂₇H₄₄N₄O₅ |
| Молекулярный вес | 504.67 |
| Хранение | Хранить в сухом прохладном месте, вдали от прямых солнечных лучей. |
N-гексановый-TyrIle-(6)-аминогексановый амид, более известный как Дигекса (Dihexa), является аналогом ангиотензина IV, проницаемым через гематоэнцефалический барьер. Дигекса классифицируется как ноотропное соединение с длительным циклическим периодом полураспада и потенциалом для повышения антидементной активности в случаях фармакологически вызванных когнитивных нарушений [1]. Текущие исследования, касающиеся Дигекса, сосредоточены на его способности улучшать восстановление периферического функционирования нервов, а также на том, как соединение взаимодействует с путем PI3K/AKT, чтобы стимулировать прокогнитивную способность ноотропа.
1) Результаты исследования показывают, что Dihexa может иметь потенциал для лечения таких заболеваний, как болезнь Альцгеймера, при которых может быть полезно усиление синаптических связей.
2) Было показано, что Dihexa улучшает когнитивные нарушения и восстанавливает память, подавляя воспаление и уменьшая потерю нейронов путем воздействия на сигнальные пути PI3K/AKT.
1) Предыдущие исследования определили способность аналогов AngIV, таких как Dihexa, увеличивать мозговой кровоток, вызывать нейропротекторные эффекты и способствовать долгосрочному потенцированию, обучению и консолидации памяти. При этом исследовательская группа McCoy et al исследовала потенциал ноотропного соединения для индукции спиногенеза и синаптогенеза. Исследование началось со сбора образцов крови из яремных вен четырехмесячных самцов крыс Sprague-Dawley. Образцы инкубировали на льду и центрифугировали для отделения сыворотки, которую затем собирали, переносили в чистые пробирки и хранили до проведения дальнейших экспериментов [2].
10 мкл каждого раствора препарата добавляли к 90 мкл образцов сыворотки крови крысы, и через определенные промежутки времени исследователи останавливали метаболизм, добавляя 1 мл ACN и уксусной кислоты, чтобы осадить белки, которые затем удаляли после ночного охлаждения с последующим центрифугированием. Супернатант отделяли, высушивали и регидратировали перед разделением и анализом ВЭЖХ. Скорость деградации препаратов измеряли, наблюдая за уменьшением площади под кривой во время удерживания препарата. Затем строили график зависимости концентрации от времени, чтобы найти кинетическую константу деградации и рассчитать период полураспада.
Микросомальный метаболизм также был изучен исследовательской группой с использованием микросом печени самцов крыс и системы регенерации НАДФН, приготовленной путем добавления 1,7 мг/мл НАДФ, 7,8 мг/мл глюкозо-6-фосфата и 6 ед/мл глюкозо-6-фосфатдегидрогеназы к 10 мл 2% бикарбоната натрия. 500 мкм раствор дигекса был приготовлен в ацетонитриле, и собранные микросомы печени были суспендированы в растворе 0,5 мг/мл 0,1 М трис-буфера при pH 7,38. Суспендированные микросомы печени были помещены на лед в предварительно охлажденные микроцентрифужных пробирках, что позволило добавить 640 мкл 0,1 М трис-буфера и 10 мкл 500 мкМ тестируемого соединения к образцу. После гибридизации rotisserie и инкубации в течение назначенного времени 500 ul каждого образца добавляли к 500 ul ледяного ацетонитрила. Затем образцы анализировали с помощью высокоэффективной жидкостной хроматографии/масс-спектрометрии для определения концентрации препарата и расчета отрицательных контрольных образцов, не содержащих микросомы печени [2].
После изучения микросомального метаболизма 24-месячные самцы крыс Sprague-Dawley весом около 290-450 граммов были использованы для поведенческих исследований. Перед поведенческими исследованиями направляющая канюля была размещена в правом полушарии и снабжена скошенным кончиком, чтобы ограничить глубину проникновения на 2,5 мм. Канюля была закреплена на черепе и запечатана толстой проволокой из нержавеющей стали. После операции животные содержались в условиях 12-часового цикла света/12-часовой темноты и находились под пристальным наблюдением и с ними осторожно обращались в течение 5 минут в день, в течение 5-6 дней после операции [2].
Первичным поведенческим исследованием был тест с водным лабиринтом, который состоял из круглого аппарата, окрашенного в черный цвет и заполненного водой на глубину 26 см. Резервуар был разделен на четыре равных квадранта, которые были определены как юго-восток, юго-запад, северо-запад и северо-восток. Скрытая платформа была случайным образом размещена в одном из четырех квадрантов и погружена на 2 см ниже поверхности воды. Точки входа в резервуар были случайным образом назначены в один из четырех углов квадранта, расположенных на севере, юге, востоке и западе, в то время как стены резервуара были покрыты пространственными подсказками разных форм и цветов. Каждый испытуемый получил инъекцию 70 нмоль гидробромида скополамина в 2 мкл искусственной спинномозговой жидкости, которая была введена в течение 20-секундного периода времени, за 20 минут до тестирования с водным лабиринтом. Затем было введено Дигекса, смешанное с 2 мкл искусственной спинномозговой жидкости, введенное за 5 минут до начала тестирования с водным лабиринтом [2].
Испытания на приобретение навыков проводились в течение 8 последовательных дней, по 5 испытаний каждый день. В первый день перед первым испытанием испытуемых помещали на платформу на 30 секунд для ознакомления. Первое испытание начиналось с того, что крысу поворачивали лицом к стене лабиринта в одной из точек входа. Затем испытуемым разрешалось плавать вокруг резервуара в течение 120 секунд, чтобы найти скрытую платформу. После того, как платформа была обнаружена, животному предоставлялся 30-секундный период отдыха перед началом следующего испытания. Если крыса не могла найти платформу, исследователи помещали крысу на платформу и давали ей 30-секундный период отдыха перед началом следующего испытания. На 9-й день после обучения приобретению навыков проводилось дополнительное испытание, в котором платформа была удалена, и крысе требовалось проплыть все 120 секунд. Это позволило исследовательской группе определить усвоенные реакции животных, подсчитав, сколько раз каждый испытуемый пересекал квадрант, где находилась платформа. Траектория плавания регистрировалась с помощью компьютерной системы слежения, которая отображала задержку плавания и пройденное расстояние [2].
2) Исследовательская группа Сана и соавторов изучила потенциал дигекса для улучшения когнитивных функций, связанных с болезнью Альцгеймера, путем воздействия на ось мозга AngIV/PI3K/AKT. Для этого исследования использовались 6-месячные самцы мышей APP/PS1 и мыши дикого типа C57. Испытуемые содержались в стандартной комнате для животных при 12-часовом световом/12-часовом темном цикле день/ночь с неограниченным доступом к пище и воде. Первая часть исследования началась со случайного разделения испытуемых на четыре группы, включая: дикий тип, APP/PS1, APP/PS1 + дигекса, вводимая в дозе 1,44 мг/кг, и APP/PS1 + дигекса, вводимая в дозе 2,88 мг/кг [1].
Вторая часть исследования началась со случайного разделения испытуемых на три группы, включая: APP/PS1, Dihexa, вводимую в дозе 2,88 мг/кг, и 2,88 мг/кг Dihexa + 0,5 мг/кг вортманнина. И Dihexa, и вортманнин были приготовлены путем растворения соединений в 10% DMSA, 40% PEG 300, 5% Tween 80 и 45% физиологическом растворе. Dihexa вводили мышам APP/PS1 внутрибрюшинно в возрасте от 6 до 9 месяцев, а 0,9% физиологический раствор вводили группе дикого типа один раз в день в течение трех месяцев.
После того, как препараты вводились испытуемым в течение 3 месяцев, они проходили тест водного лабиринта Морриса. Круглая черная ванна была наполнена водой и разделена на четыре равные области, обозначенные как север (С), юг (Ю), восток (В) и запад (З). Небольшая платформа была погружена на 1 см ниже поверхности воды в центре северо-восточного квадранта резервуара. Каждая мышь проходила 4 испытания в день в течение 5 последовательных дней. Им давали 60 секунд, чтобы обыскать резервуар в поисках платформы, и в конце каждого сеанса их помещали на платформу и оставались там в течение 30 секунд. На 6-й день платформу убирали, и исследователи регистрировали количество раз, когда отдельные мыши пересекали квадрант, где ранее находилась платформа, в течение 60 секунд [1].
После того, как все необходимые данные были собраны для теста водного лабиринта Морриса, мышей умертвили, а мозговую ткань препарировали. Полученную мозговую ткань взвешивали и добавляли PBS, чтобы обеспечить соотношение веса (г) к объему (мл) 1:9. Образцы гомогенизировали, чтобы можно было центрифугировать супернатант, после чего измеряли уровни AngIV, TNF-альфа, IL-10 и IL-1-бета с помощью чувствительного и специфического анализа ELISA. После перфузии мозг фиксировали в течение 48 часов и заливали в парафин. Затем парафиновые срезы толщиной 4 мкм обезвоживали, окрашивали метиленовым синим и промывали дистиллированной водой, после чего проводили повторную дегидратацию и промывку срезов для подготовки к окрашиванию по Нисслю. Все положительно окрашенные клетки подсчитывались, и «% положительных по Нисслю нейронов» рассчитывался путем деления числа положительных нейронов на общее число нейронов в каждой клетке [1].
Наконец, образцы были подготовлены для анализа вестерн-блоттинга путем лизиса гомогенатов мозга на льду в течение 300 минут в 100 мл буфера для лизиса, состоящего из 120 мМ NaCL, 40 мМ Трис (pH 8) и 0,1% NP40, с последующим центрифугированием. Кроме того, для определения концентрации белка использовался анализ бицинхониновой кислоты (BCA); 30 мкг белка было разделено с помощью 10% SDS-PAGE и электроблоттинга на мембранах из поливинилидендифторида (PVDF). Мембраны были заблокированы в 5% обезжиренном молоке и инкубированы в течение ночи при 4 градусах Цельсия с первичными антителами. За этой процедурой последовала инкубация мембран со вторичными антителами при комнатной температуре в течение 2 часов [1].
1) Целью исследовательской группы Маккоя и соавторов было найти аналог AngIV, способный проникать через гематоэнцефалический барьер и вызывать прокогнитивную активность путем улучшения метаболической стабильности три- и тетрапептидных N-концев аналогов AngIV. Различные структурные изменения были сделаны на N-конце, такие как замена D-норлейцина на L-норлейцин, N-ацетилирование норлейцина и замена норлейцина на ГАМК с целью улучшения общей стабильности путем снижения восприимчивости к аминопептидазам.
Соединения инкубировали в присутствии образцов сыворотки крови крысы, и все полученные инкубаты анализировали с помощью ВЭЖХ для определения скорости их метаболической активности. Результаты показали, что AngIV имел очень короткий период полураспада, менее 2 минут, однако каждое из модифицированных N-концевых соединений, таких как Dihexa, демонстрировало более продолжительные периоды полураспада по сравнению с исходным соединением. Эти результаты свидетельствуют о том, что снижение деградации N-конца приводит к улучшению метаболической стабильности [2].
Дигекса также помогает улучшить метаболическую стабильность благодаря высокому уровню биодоступности для распределения в тканях. В дополнение к его биодоступности и длительному периоду полувыведения, микросомы печени крыс были исследованы в качестве дополнительного измерения эффектов Дигекса на скорость метаболизма. Первоначальные результаты показали, что метаболизм фазы I ноотропа был очень низким с внутренним клиренсом 2,72 мкл/мин/мг, а также средним периодом полувыведения приблизительно 510 минут. Скорость клиренса Дигекса сравнивали с аналогичными ноотропными соединениями, такими как пироксикам, верапамил и 7-этоксикумарин [2].
С точки зрения поведенческого тестирования, Dihexa оценивали на предмет его способности устранять дефициты, вызванные введением скополамина, наблюдаемые в результатах испытуемых в тесте с водным лабиринтом. Первоначальное испытание было проведено для проверки нейропротекторного и когнитивного действия Dihexa; собранные результаты показали, что как высокие, так и низкие дозы ноотропа значительно повышают результаты в тесте с водным лабиринтом по сравнению с крысами, которым вводили только скополамин. Важно отметить, что во все дни тестирования группа, получавшая более высокую дозу Dihexa, демонстрировала результаты, которые были неотличимы от контрольной группы, которой вводили только соединения-носители [2].
Кроме того, исследовательская группа стремилась определить, были ли какие-либо существенные изменения в результатах в зависимости от того, вводился ли ноотроп через канюлю, внутрибрюшинную инъекцию или пероральное введение. Для всех трех методов введения наблюдалась четкая зависимость между дозой и реакцией между результатами в тесте водного лабиринта и введением Дигексы. При этом высокие дозы ноотропа, измеренные на уровне 0,5 мг/кг/день для внутрибрюшинного введения и 2,0 мг/кг/день для перорального введения, привели к результатам, которые были неотличимы от контрольной группы и значительно лучше, чем у животных, получавших только скополамин [2].
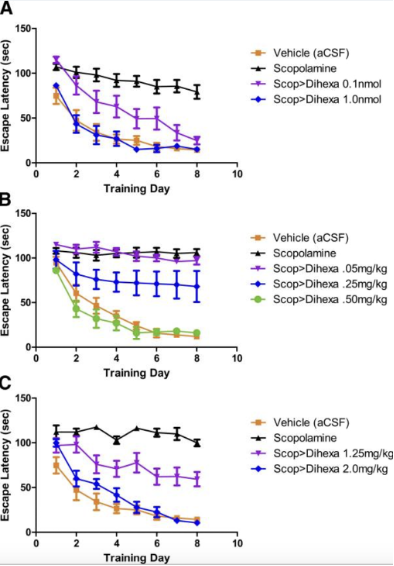
Рисунок 1: Изменения дефицита обучения, зависящего от скополамина, в ответ на лечение Дигекса. A) Крысы, которым предварительно вводили 70 нмоль скополамина и которым вводили Дигекса через канюлю. B) Крысы, которым предварительно вводили 70 нмоль скополамина и которым вводили Дигекса посредством внутрибрюшинной инъекции. C) Крысы, которым предварительно вводили 70 нмоль скополамина и которым вводили Дигекса пероральным способом.
После 8 дней обучения приобретению навыков начался 9-й день испытаний, когда платформа была удалена от аппарата, и крысы должны были плыть в течение полных 120 секунд, в то время как исследовательская группа регистрировала, сколько раз каждая крыса проходила в квадрант, где ранее находилась платформа. Испытуемые, которым вводили самую высокую дозу Дигексы, проводили больше всего времени в целевом квадранте по сравнению с группами животных, которым вводили только скополамин. При этом результаты показали, что между введением ноотропа и зарегистрированными латентностями побега существовала дозозависимая связь [2].
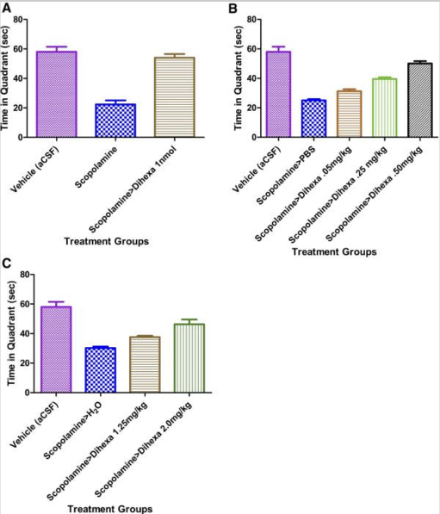
Рисунок 2: Время, проведенное в целевом квадранте испытуемыми в каждой экспериментальной группе лечения. A) мыши, которым вводили Дигекса через канюлю, B) мыши, которым вводили Дигекса посредством внутрибрюшинной инъекции, и C) мыши, которым вводили Дигекса пероральным способом доставки.
2) Для того чтобы определить, участвует ли AngIV в развитии болезни Альцгеймера, исследовательская группа Сана и др. определила базовые уровни AngIV как у мышей дикого типа, так и у мышей APP/PS1. По сравнению с мышами дикого типа, у мышей APP/PS1 были обнаружены значительно более низкие уровни AngIV в мозге. При этом крысам вводили Дигекса в дозах 1,44 мг/кг и 2,88 мг/кг, чтобы увидеть, как уровни AngIV изменяются в ответ на ноотропное соединение. Результаты показали, что обе дозы Дигекса повышали уровни AngIV в мозге мышей APP/PS1, причем доза 2,88 мг/кг повышала эти уровни почти до того же уровня у мышей дикого типа. Эти результаты свидетельствуют о том, что уровни AngIV в мозге потенциально играют роль в развитии болезни Альцгеймера [1].
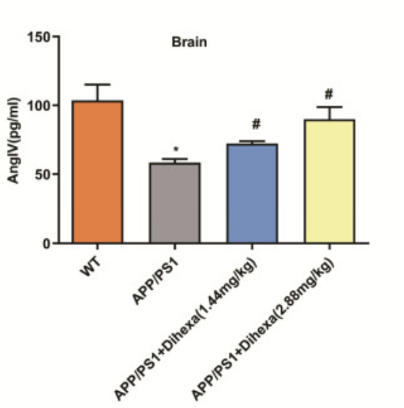
Рисунок 3: Средние уровни AngIV в мозге в четырех различных экспериментальных группах, включенных в первую часть исследования.
В дополнение к уровням AngIV в мозге, испытуемые прошли тест водного лабиринта Морриса, чтобы измерить когнитивные способности мышей APP/PS1 при введении Дигексы. С 1-го по 5-й день эксперимента было обнаружено, что латентность побега значительно снизилась, однако латентность побега у мышей APP/PS1 была выше, чем у мышей дикого типа. При этом обе дозы ноотропа 1,44 мг/кг и 2,88 мг/кг были успешными в снижении латентности побега в различной степени, причем этот эффект был наиболее выражен на 4-й и 5-й дни эксперимента. В целом, исследователи обнаружили, что мыши APP/PS1, получавшие Дигексу, продемонстрировали значительно лучшие результаты по сравнению с контрольными мышами при оценке количества пересечений платформы. Эти результаты указывают на то, что лечение Дигексой улучшает когнитивные способности у мышей APP/PS1 [1].
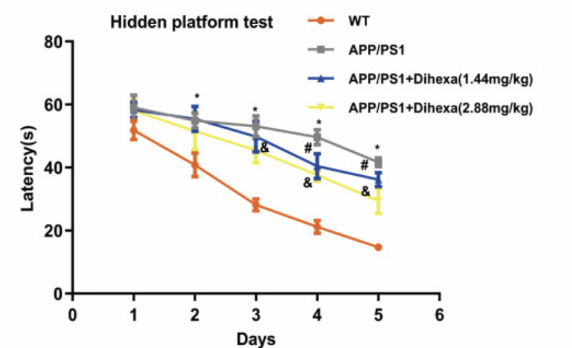
Рисунок 4: Изменения латентности побега в четырех различных экспериментальных группах, включенных в первую часть исследования.
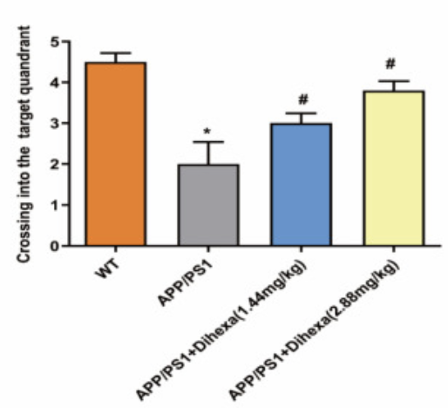
Рисунок 5: Среднее количество переходов в целевой квадрант в каждой из четырех различных экспериментальных групп, включенных в первую часть исследования.
После теста с водным лабиринтом Морриса для наблюдения за количеством присутствующих положительных нейронных клеток использовалось окрашивание по Нисслю. По сравнению с мышами дикого типа, мыши APP/PS1 испытывали значительную синаптическую потерю, а также сокращение количества нейронных клеток в коре головного мозга. При лечении как дозой 1,44 мг/кг, так и дозой 2,88 мг/кг дигекса, мыши APP/PS1 испытывали увеличение количества нейронных клеток, присутствующих в коре головного мозга. Основываясь на результатах окрашивания по Нисслю, исследовательская группа смогла сделать вывод, что лечение дигекса ослабило скорость потери нейронов в мозге мышей APP/PS1 [1].
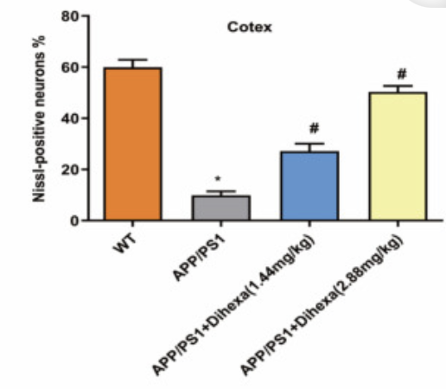
Рисунок 6: Процент нейронов, положительных по Нисслю, в каждой из четырех различных экспериментальных групп, включенных в первую часть исследования.
Кроме того, для того чтобы изучить механизм действия нейронального апоптоза, исследовательская группа оценила уровни нейровоспаления и глиальной активации, обнаружив уровни IL-1-бета, IL-10 и TNF-альфа в мозге. Исходные измерения показали, что уровни TNF-альфа и IL-1-бета в группе мышей APP/PS1 были намного выше, чем в группе мышей дикого типа. Однако, когда мышей лечили Дигексой, было обнаружено, что уровни как IL-1-бета, так и TNF-альфа значительно снизились. С другой стороны, исходные измерения показали, что уровни IL-10 у мышей APP/PS1 были значительно снижены по сравнению с мышами дикого типа, а при лечении Дигексой у этих мышей наблюдалось повышение уровней IL-10. Эти результаты свидетельствуют о том, что ноотропное соединение оказывает нейропротекторное действие на нервные клетки в мозге, поврежденные воспалительными факторами [1].
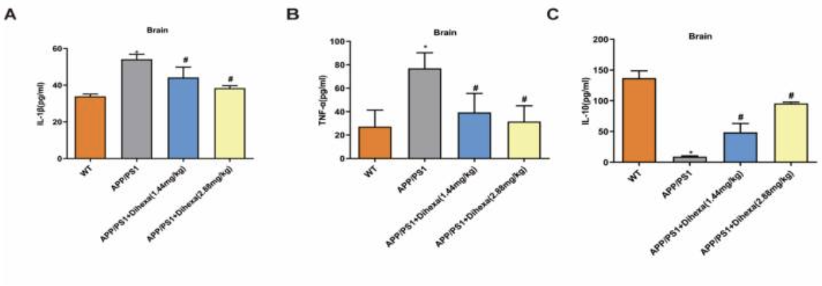
Рисунок 7: Изменения уровней A) ИЛ-1-бета, B) ФНО-альфа и C) ИЛ-10 в каждой из четырех различных экспериментальных групп, включенных в первую часть исследования.
Важно отметить, что исследовательская группа продвинула свои экспериментальные процедуры на шаг дальше, чтобы определить связь между Dihexa и сигнальным путем PI3K/AKT, вортманнин, ингибитор PI3K, вводился мышам внутрижелудочно для определения количества нейронных клеток и присутствующих воспалительных факторов. Было обнаружено, что введение вортманнина значительно отменяет экспрессию PI3K и AKT, а также антиапоптотические и противовоспалительные эффекты Dihexa, что приводит к уменьшению количества нейронных клеток, присутствующих в коре, и уровней IL-10, а также повышению уровней TNF-альфа и IL-1-бета [1].
[1] Sun X, Deng Y, Fu X, Wang S, Duan R, Zhang Y. AngIV-Analog Dihexa Rescues Cognitive Impairment and Recovers Memory in the APP/PS1 Mouse via the PI3K/AKT Signaling Pathway. Brain Sci. 2021 Nov 11;11(11):1487. doi: 10.3390/brainsci11111487. PMID: 34827486; PMCID: PMC8615599.
[2] McCoy AT, Benoist CC, Wright JW, Kawas LH, Bule-Ghogare JM, Zhu M, Appleyard SM, Wayman GA, Harding JW. Evaluation of metabolically stabilized angiotensin IV analogs as procognitive/antidementia agents. J Pharmacol Exp Ther. 2013 Jan;344(1):141-54. doi: 10.1124/jpet.112.199497. Epub 2012 Oct 10. PMID: 23055539; PMCID: PMC3533412.
© 2026 АнкеБио. Magazin nootropov Biohaker
127299, г. Москва, ул. Шоссе Энтузиастов, 56 стр. 32
+7 (495) 227-22-05
+7 (985) 227-22-05
Email: ankebiorus@gmail.com
Вы нашли для себя препарат, который хотите
приобрести по низкой цене и с гарантией
безопасной доставки?
Звоните нам
Мы Вам обязательно ответим!